


ABSTRACT
Background and Aim: Benzimidazoles (BZ) remain the cornerstone of soil-transmitted helminth control programs; however, the emergence of resistance-associated mutations in the β-tubulin gene threatens their long-term effectiveness. Information on BZ resistance markers in hookworms from Southeast Asia remains limited. This study investigated the occurrence of β-tubulin isotype 1 mutations in hookworms from the Philippines and Indonesia and evaluated the potential effects of detected variants on BZ binding and tubulin dimerization using in silico analyses.
Materials and Methods: Eighteen archived hookworm-positive fecal samples collected from the Philippines (n = 13) and Indonesia (n = 5) were analyzed. Molecular speciation was performed using species-specific genetic markers, followed by amplification and sequencing of the β-tubulin isotype 1 gene regions that encompass known BZ resistance codons. Identified variants were subjected to in silico molecular docking, molecular dynamics simulations, binding free-energy calculations, and protein–protein interaction analyses to assess their effects on drug binding and tubulin dimer formation.
Results: Fifteen samples were identified as Necator americanus and three as Ancylostoma ceylanicum. No canonical BZ resistance-associated mutations were detected at codons 134, 167, 198, or 200. However, two A. ceylanicum isolates from the Philippines carried a novel V236I substitution resulting from a GTC→ATC nucleotide change. Molecular docking demonstrated minimal alterations in docking poses, whereas molecular dynamics simulations revealed a marked reduction in BZ-binding favorability in the mutant protein. The V236I variant exhibited substantially lower MM-GBSA binding free-energy with albendazole than the wild-type protein (−13.34 vs. −33.42 kcal/mol), accompanied by reduced hydrogen-bond formation. Structural analyses suggested that the bulkier isoleucine side chain may introduce steric hindrance within the binding region, thereby weakening ligand–protein interactions. In contrast, the mutation produced negligible effects on tubulin dimer stability, binding affinity, and conformational dynamics.
Conclusion: A novel V236I substitution was identified in A. ceylanicum from Southeast Asia and was predicted to reduce BZ-binding efficiency through steric and energetic effects while preserving tubulin dimer integrity. These findings highlight V236I as a potential BZ resistance-associated variant and emphasize the need for continued molecular surveillance and experimental validation of emerging resistance markers in hookworm populations.
Keywords: albendazole, Ancylostoma ceylanicum, benzimidazole resistance, β-tubulin, hookworm, molecular dynamics, soil-transmitted helminths, Southeast Asia.
INTRODUCTION
Benzimidazole (BZ) resistance is an emerging threat to public and veterinary health [1]. BZ anthelmintics, such as albendazole (ABZ), mebendazole, and fenbendazole (FBZ), remain the cornerstone of clinical treatment against helminth infections and are widely used in population-based infection control and prevention programs in both animals and humans [2, 3]. The core structure of BZ drugs consists of fused benzene and imidazole rings with varying moieties at positions 2, 5, or 6 that enhance absorption and binding affinity to target helminth proteins [4]. These compounds disrupt microtubule dynamics by binding to β-tubulin, thereby impairing essential cytoskeletal functions and glucose transport pathways that are critical for parasite survival [5, 6]. Helminths can evade the anthelmintic activity of BZs through alterations in the amino acid (aa) composition of the β-tubulin protein. Single-nucleotide polymorphisms (SNPs) in the β-tubulin isotype 1 gene that result in key aa substitutions are recognized as the primary molecular mechanism underlying BZ resistance in numerous helminth species of veterinary and public health importance [7, 8]. The most commonly reported substitutions include F167Y (TTC, TTT → TAC, TAT), E198A (GAG, GAA → GCG, GCA), and F200Y (TTC, TTT → TAC, TAT). Experimental studies have confirmed the roles of these mutations in conferring BZ resistance [9–11]. These resistance-associated mutations have been reported in soil-transmitted helminths (STHs), including Ascaris lumbricoides, Trichuris trichiura, and several hookworm species from different regions of the world [12, 13].
The emergence of BZ resistance is of particular concern in hookworms, given the increasing number of reports documenting resistance-associated mutations in both animal and human parasites. This issue has been extensively documented in Ancylostoma caninum, a canine hookworm species that is frequently subjected to repeated anthelmintic treatments in the United States and Canada [14, 15]. Several studies have reported the widespread occurrence of the F167Y and Q134H (CAA → CAT) mutations in North America [16–18]. More recently, the F167Y mutation has been reported in A. caninum and Uncinaria stenocephala from Australia and New Zealand [19]. Among human hookworms, resistance-associated mutations have also been identified in Necator americanus populations from Ghana [13], Brazil [20], Mozambique [21], Haiti, Kenya, and Panama [12]. The increasing geographic distribution of these mutations highlights the potential for reduced efficacy of current BZ-based control strategies. Their occurrence warrants continuous monitoring because the establishment and spread of resistant hookworm populations may compromise ongoing control and elimination programs in both humans and animals [22, 23]. This concern is particularly relevant in STH-endemic countries where preventive chemotherapy has been implemented over prolonged periods [24].
Despite the growing global evidence of BZ resistance, information regarding resistance-associated mutations in hookworms from Southeast Asia remains scarce. Reports from several countries in the region have suggested a decline in the efficacy of BZ-based preventive chemotherapy programs, raising concerns about the potential emergence of drug-resistant parasite populations [25, 26]. Southeast Asia remains highly endemic for STH infections, including hookworm infections, and many countries continue to rely heavily on mass drug administration and other drug-based control interventions as the primary means of disease management [27–30]. Consequently, the emergence and dissemination of resistance-associated mutations could substantially undermine regional control efforts and threaten the long-term sustainability of existing intervention programs.
A knowledge gap currently exists regarding the molecular epidemiology of BZ resistance markers in hookworms circulating in Southeast Asia. Most available studies have focused on resistance-associated mutations in Africa, the Americas, and Oceania, whereas molecular surveillance data from Southeast Asian hookworm populations remain limited. Furthermore, little is known about the occurrence of novel variants outside the canonical resistance codons or their potential functional consequences on BZ–β-tubulin interactions. The identification and characterization of such variants are essential for improving resistance surveillance programs and for understanding the evolutionary mechanisms that may contribute to reduced drug susceptibility in hookworms.
Therefore, the present study aimed to investigate the occurrence of mutations in the β-tubulin isotype 1 gene of hookworms obtained from the Philippines and Indonesia and to evaluate the potential functional consequences of detected variants through comprehensive in silico analyses. Specifically, this study sought to identify known and novel BZ resistance-associated mutations and to assess their potential effects on BZ binding and tubulin dimerization. By combining molecular characterization with structural and dynamic computational analyses, this study provides new insights into emerging genetic variants that may contribute to reduced BZ susceptibility in hookworm populations from Southeast Asia.
MATERIALS AND METHODS
Ethical approval
This study utilized only archived, de-identified leftover fecal samples previously collected from hookworm-positive individuals in Leyte Province, Philippines (adults), and Southwest Sumba Regency, Indonesia (school-aged children). No new participant recruitment, field sampling, or collection of identifiable personal data occurred.
Ethical approval for the original sample collection and use in the Philippines was granted by the University of the Philippines Manila Research Ethics Board (Approval Code: 2017-369-01). In Indonesia, approval was obtained from the Ethical committee of the School of Medicine and Health Sciences, Atma Jaya Catholic University of Indonesia (Approval Code: 25/08/KEP-FKIKUAJ/2023). The current study, which involved molecular speciation, β-tubulin gene amplification/sequencing, and in silico analyses of these archived specimens, received separate ethical approval from the Khon Kaen University Center for Ethics in Human Research (Approval Code: HE681112, March 6, 2025).
Because the study used only de-identified leftover diagnostic specimens, the ethics committees waived the requirement for additional informed consent. All procedures were conducted in accordance with the ethical principles of the Declaration of Helsinki (as revised in 2013), the International Conference on Harmonisation Good Clinical Practice guidelines, and applicable national regulations in the Philippines, Indonesia, and Thailand governing the secondary use of archived human biological materials for research. Strict data confidentiality and privacy protections were maintained throughout the study; no personally identifiable information was accessed or stored by the research team.
Study period and location
The study was conducted at the Department of Tropical Medicine, Faculty of Medicine, Khon Kaen University, Khon Kaen, Thailand, from November 2023 to July 2025. Samples from the Philippines were originally collected from adults residing in Leyte Province, whereas samples from Indonesia were collected from school-aged children in Southwest Sumba Regency. Molecular laboratory procedures, sequence analyses, and computational evaluations were performed at Khon Kaen University during the study period.
Study design
This study employed a retrospective molecular epidemiological design using archived hookworm-positive fecal samples. No additional participant recruitment or field sampling was conducted because only available leftover or archived specimens were utilized. The study involved molecular species identification, amplification and sequencing of target regions of the β-tubulin isotype 1 gene, variant detection, and subsequent in silico analyses to evaluate the potential effects of identified mutations on BZ-binding and tubulin dimerization.
Amplification of A. Ceylanicum cytochrome c oxidase subunit I (cox1) and N. Americanus ITS1, 5.8S, and ITS2 genes
DNA was extracted from hookworm-positive fecal samples using the TIANamp Stool DNA Kit (Tiangen Biotech Co., Ltd., Beijing, China) according to the manufacturer's instructions. A segment of the A. ceylanicum cox1 gene was amplified using conventional polymerase chain reaction (PCR). Primers were designed using Primer-BLAST (https://www.ncbi.nlm.nih.gov/tools/primer-blast/) based on the available A. ceylanicum cox1 sequence in GenBank (GeneID: 33194677). The primer sequences were AceyCox1F (5′-TGTTTGGATCTTTGGGG ATGGT-3′) and AceyCox1R (5′-ACGGGGAAAACCATGTAAACCT-3′).
The 25 μL PCR reaction mixture consisted of 12.5 μL RealStart DNA Polymerase Premix (Yeastern Biotech Co., Ltd., Taipei, Taiwan), 1 μL (10 μM) each of forward and reverse primers, 6.5 μL diethyl pyrocarbonate (DEPC)-treated H₂O, and 4 μL DNA template. PCR amplification was performed under the following conditions: initial denaturation at 94°C for 15 min, followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 58°C for 30 s, and extension at 72°C for 30 s, with a final extension at 72°C for 7 min. PCR amplicons were visualized through electrophoresis on 1.5% agarose gels at 75 V for 30 min.
The Internal transcribed spacer (ITS)1, 5.8S, and ITS2 regions of N. americanus were amplified using primers previously reported by Diawara et al. [12]: RTHW1F (5′-GATGAGCATTGCWTGAATGCCG-3′) and RTHW1R (5′-GCAAGTRCCGTTCGACAAACAG-3′). The 25 μL PCR mixture consisted of 12.5 μL RealStart DNA Polymerase Premix (Yeastern Biotech Co., Ltd., Taipei, Taiwan), 1 μL (10 μM) each of forward and reverse primers, 6.5 μL DEPC-treated H₂O, and 4 μL DNA template. PCR conditions included an initial denaturation at 94°C for 15 min, followed by 35 cycles of 94°C for 30 s, 62°C for 30 s, and 72°C for 30 s, with a final extension at 72°C for 7 min. PCR amplicons were visualized by electrophoresis on a 1.5% agarose gel at 75 V for 30 min.
Amplification and sequencing of the β-tubulin isotype 1 gene
Specific regions containing BZ resistance-associated codons (aa positions 134, 167, 198, and 200) were amplified using two previously published protocols (Table 1). For samples confirmed as A. ceylanicum, a nested PCR protocol reported by Medeiros et al. [31] was followed with minor modifications. These modifications include changes in the constitution of the PCR mix due to and thermocycler conditions due to the use of a commercially available kit.
For the first PCR round, the 25 μL reaction mixture contained 12.5 μL RealStart DNA Polymerase Premix (Yeastern Biotech Co., Ltd., Taipei, Taiwan), 1 μL (10 μM) each of primers (Fa167Ay and Ra167Ay for aa134/167; Fa198/200Ay and Ra198/200Ay for aa198/200), 6.5 μL DEPC-treated H₂O, and 4 μL DNA template. For the second round, the reaction mixture consisted of 12.5 μL RealStart DNA Polymerase Premix, 1 μL (10 μM) each of primers (Fc167Ay and Rc167Ay for aa134/167; Fc198/200Ay and Rc198/200Ay for aa198/200), 6.5 μL DEPC-treated H₂O, and 4 μL first-round PCR product. The primer sequences and amplicon size for this experiment is shown in Table 1 [31].
PCR conditions for both rounds consisted of initial denaturation at 94°C for 15 min, followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 58°C for 45 s, and extension at 72°C for 45 s, with a final extension at 72°C for 7 min. Amplicons were visualized using 1.5% agarose gel electrophoresis at 75 V for 30 min.
For samples positive for N. americanus, a conventional PCR protocol based on primers reported by Diawara et al. [12] was employed (Table 1). The 25 μL PCR mixture consisted of 12.5 μL RealStart DNA Polymerase Premix (Yeastern Biotech Co., Ltd., Taipei, Taiwan), 1 μL (10 μM) each of primers (HwTub134/167F and HwTub134/167R for aa134/167; HwTub198/200F and HwTub198/200R for aa198/200), 6.5 μL DEPC-treated H₂O, and 4 μL DNA template.
For the aa134/167 fragment, PCR conditions included initial denaturation at 94°C for 15 min, followed by 35 cycles at 94°C for 30 s, 62°C for 45 s, and 72°C for 45 s, with a final extension at 72°C for 7 min. For the aa198/200 fragment, amplification was performed under the same cycling conditions, except with an annealing temperature of 58°C. Amplicons were visualized using 1.5% agarose gel electrophoresis at 75 V for 30 min.
Samples exhibiting clear amplification bands were submitted for next-generation sequencing (NGS) to U2Bio Co., Ltd. (Bangkok, Thailand) through its Fast NGS service using the Illumina sequencing platform.
| Species | Target region | PCR round | Primer | Sequence (5′–3′) | Amplicon size (bp) | Reference |
|---|---|---|---|---|---|---|
| A. ceylanicum | aa134/167 | External | Fa167Ay | TGAGCTCGTCGATAACGTCC | 415 | [31] |
| - | - | - | Ra167Ay | TGAGGTCATCCCCAGTTTGAC | - | - |
| - | aa198/200 | External | Fa198/200Ay | TGTTCCTAAAAAGGGGTCGGG | 424 | [31] |
| - | - | - | Ra198/200Ay | AAGCGAAGGCAGGTAGTGAC | - | - |
| - | aa134/167 | Internal | Fc167Ay | CAGGTATTTCGCAACCGTGC | 306 | [31] |
| - | - | - | Rc167Ay | AGGAACACGACCAGCGTTT | - | - |
| - | aa198/200 | Internal | Fc198/200Ay | GCAGTCCACGTTCCTGCTTA | 308 | [31] |
| - | - | - | Rc198/200Ay | ACCGGACATTGTTACAGACACT | - | - |
| N. americanus | aa134/167 | Conventional | HwTub134/167F | GTGACTGTCTCCAGGTAATTCG | 220 | [12] |
| - | - | - | HwTub134/167R | CTATAACGTACCTTTGGCGAGGG | - | - |
| - | aa198/200 | Conventional | HwTub198/200F | TTTCCGACACTGTGGTTGAG | 185 | [12] |
| - | - | - | HwTub198/200R | GAGTTCGTTACTAGCCAGCTCACC | - | - |
Table 1. Primers used for amplification of the β-tubulin isotype 1 gene in different hookworm species.
| Species | Target region | PCR round | Primer | Sequence (5′–3′) | Amplicon size (bp) | Reference |
|---|---|---|---|---|---|---|
| A. ceylanicum | aa134/167 | External | Fa167Ay | TGAGCTCGTCGATAACGTCC | 415 | [31] |
| - | - | - | Ra167Ay | TGAGGTCATCCCCAGTTTGAC | - | - |
| - | aa198/200 | External | Fa198/200Ay | TGTTCCTAAAAAGGGGTCGGG | 424 | [31] |
| - | - | - | Ra198/200Ay | AAGCGAAGGCAGGTAGTGAC | - | - |
| - | aa134/167 | Internal | Fc167Ay | CAGGTATTTCGCAACCGTGC | 306 | [31] |
| - | - | - | Rc167Ay | AGGAACACGACCAGCGTTT | - | - |
| - | aa198/200 | Internal | Fc198/200Ay | GCAGTCCACGTTCCTGCTTA | 308 | [31] |
| - | - | - | Rc198/200Ay | ACCGGACATTGTTACAGACACT | - | - |
| N. americanus | aa134/167 | Conventional | HwTub134/167F | GTGACTGTCTCCAGGTAATTCG | 220 | [12] |
| - | - | - | HwTub134/167R | CTATAACGTACCTTTGGCGAGGG | - | - |
| - | aa198/200 | Conventional | HwTub198/200F | TTTCCGACACTGTGGTTGAG | 185 | [12] |
| - | - | - | HwTub198/200R | GAGTTCGTTACTAGCCAGCTCACC | - | - |
Note Aa = Amino acid, PCR = Polymerase chain reaction, bp = Base pairs.
Analysis of amplicon sequences
Raw FastQ files were uploaded to the analysis server, and the corresponding hookworm β-tubulin reference genes (WormBase ParaSite Gene ID: Acey_s0035 for A. ceylanicum and WormBase ParaSite Gene ID: NAME_02123 for N. americanus) were indexed as references. Sequence quality was initially assessed using the FastQC tool (https://toolshed.g2.bx.psu.edu/view/devteam/fastqc), with particular attention given to per-base sequence quality scores and adapter content.
Sequences with Phred quality scores below 30 and adapter sequences were removed using TrimGalore. Following trimming, a second FastQC analysis was performed to verify sequence quality. Reads were mapped to the corresponding reference genes using BWA-MEM2, generating BAM files. Variant calling was performed using the bcftools mpileup and bcftools call utilities. The bcftools count function was used to determine the numbers of samples, SNPs, insertions/deletions, multiple-nucleotide polymorphisms, and total variant sites.
To ensure high-confidence variant identification, a variant call format (VCF) filter was applied with thresholds of quality score >200 and average sequencing depth ≥30. Filtered variants were subsequently used to generate consensus sequences using bcftools consensus. Variant effects on aa substitutions were confirmed using the WormBase Parasite Variant Effect Predictor (https://parasite.wormbase.org/Multi/ Tools/VEP).
Molecular docking and dynamics with BZ drugs
The mutation identified through sequencing was introduced into the A. ceylanicum β-tubulin isotype 1 protein sequence (UniProt ID: A0A0D6MC88). The three-dimensional structure of the mutant protein was generated using SwissModel (https://swissmodel.expasy.org/). Structural quality was evaluated using ProSA, Verify3D, and Ramachandran plot analysis (Supplementary Figure 5).
The modeled mutant protein was docked with ABZ and FBZ. These compounds were selected to facilitate direct comparison with our previous findings [32]. Molecular docking was performed using AutoDock Vina [33] implemented in PyRx Virtual Screening Tool version 0.8. Appropriate protonation states and polar hydrogen atoms were assigned before docking, and ligand molecules were energy-minimized.
Docking simulations were performed using a 25 Å search grid centered on aa position 200 (X = 15.6, Y = 57.5, Z = 33.1) [34]. Docking poses with a root-mean-square deviation (RMSD) of 0 were selected and visualized using BIOVIA Discovery Studio Visualizer version 21.1.0.20298.
The docked β-tubulin–BZ complexes were subsequently subjected to molecular dynamics simulations to evaluate dynamic ligand–protein interactions. The CHARMM General Force Field server was used to generate ligand topologies, whereas the CHARMM36 force field was used for protein topology generation. The complexes were solvated in cubic simulation boxes with 10 Å buffers, filled with transferable intermolecular potential with 3-point water molecules, neutralized with Na⁺ and Cl⁻ ions, and supplemented with 0.15 M NaCl.
Energy minimization was performed using 50,000 steepest-descent steps, followed by equilibration at 300 K and 1 bar using the number-of-particles, volume, and temperature (NVT) and number-of-particles, pressure, and temperature (NPT) ensembles for 100 ps each. A 100 ns production MD simulation was subsequently performed, generating 1,000 trajectory frames. RMSD, root mean square fluctuation (RMSF), solvent-accessible surface area (SASA), and hydrogen-bond analyses were conducted to evaluate ligand–protein interactions. System equilibration was achieved after approximately 80 ns of simulation.
Absolute binding free energies were estimated using molecular mechanics generalized Born surface area (MM-GBSA) and molecular mechanics Poisson–Boltzmann surface area (MM-PBSA) analyses based on the final 20 ns of equilibrated trajectories. Per-residue free-energy decomposition analyses were also performed using MM-GBSA. Results were compared with previously reported molecular dynamics analyses of the Q134H and E198K mutations [32].
Effect on tubulin dimerization
In this in silico investigation, A. ceylanicum α-tubulin (UniProt ID: A0A016VPU0) and β-tubulin (UniProt ID: A0A0D6MC88) proteins were used as structural models [34]. The identified mutation was encoded within the β-tubulin monomer, and the α/β-tubulin dimer structure was modeled using AlphaFold 3 [35].
Guanosine triphosphate (GTP) molecules were incorporated to simulate their biological role in microtubule dynamic instability mediated through GTP binding and hydrolysis [36]. Structural quality assessments included analysis of the predicted local distance difference test (pLDDT), inter-chain predicted template modeling score (ipTM), predicted template modeling score (pTM), and predicted aligned error plots. The generated structures were refined using the GalaxyRefineComplex server [37] and subsequently assessed using ProSA, Verify3D, and Ramachandran plot analysis (Supplementary Figure 6).
The effects of the β-tubulin mutation on dimerization were evaluated through several complementary approaches. Binding free energies between α- and β-subunits were estimated using HawkDock and MM-GBSA analyses [38]. Individual aa contributions to dimer stability were also examined. Binding affinity (ΔG) and dissociation constant (Kd) values were estimated using the PRODIGY server (https://wenmr.science.uu.nl/ prodigy/manual).
Molecular dynamics simulations of tubulin dimers were performed using the WebGro server with the GROMOS96 43a1 force field. Systems were solvated in simple point-charge water, neutralized with 0.15 M NaCl, and subjected to energy minimization over 5,000 steepest-descent steps. Following equilibration under Constant NVT and Constant NPT conditions at 300 K and 1.0 bar, 50 ns production simulations were performed with structures saved every 50 ps. RMSD, RMSF, radius of gyration, and SASA analyses were conducted. Results were compared with our previously published molecular dynamics analyses of the Q134H and E198K mutants [34].
RESULTS
Mutation in the β-tubulin gene
This study involved hookworm-positive fecal samples obtained from the Philippines (n = 13) and Indonesia (n = 5). Among the samples from the Philippines, three were identified as A. ceylanicum and 10 were identified as N. americanus through amplification of the cox1 and ITS1, 5.8S, and ITS2 gene regions, respectively (Supplementary Figure 1). The samples were subsequently subjected to amplification and sequencing of specific regions of the β-tubulin isotype 1 gene associated with BZ resistance.
For each sample, two target regions were amplified and sequenced: aa134/167, encompassing aa positions 134 and 167, and aa198/200, encompassing aa positions 198 and 200 (Figure 1). One A. ceylanicum sample failed amplification and sequencing of the aa134/167 fragment (Figure 1A and 1B). Among N. americanus samples, eight failed amplification and sequencing of the aa134/167 fragment, whereas only one sample failed amplification of the aa198/200 fragment (Figure 1C).
Sequence analysis of the β-tubulin gene revealed that no SNPs resulted in aa substitutions at the canonical BZ resistance-associated codons 134, 167, 198, and 200 in either hookworm species. However, analysis of the aa198/200 fragment identified a novel mutation at aa position 236. At this position, the wild-type valine residue was replaced by isoleucine through a single-nucleotide substitution (GTC → ATC) (Figure 2B). This V236I mutation was detected in two A. ceylanicum isolates from the Philippines. The mutation exhibited allele frequencies of 43% and 37% in the respective samples.
The mapped sequence alignments (BAM files) and filtered VCF files are presented in Supplementary Figures 2 and 3. Additional synonymous nucleotide substitutions were also observed in the filtered VCF files (Supplementary Figure 3).
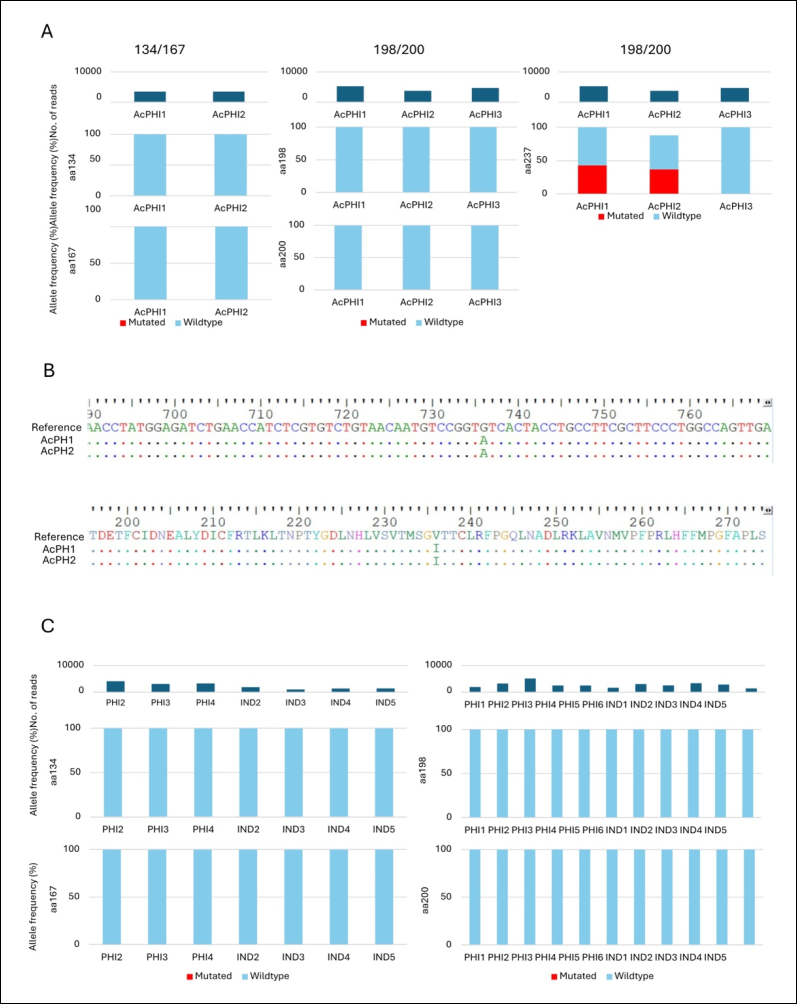
Figure 1. Sequencing analysis of the β-tubulin isotype 1 gene in hookworms. Two regions of the hookworm β-tubulin isotype 1 gene containing known BZ resistance-associated codons were amplified and sequenced (A and C). No mutations were detected at amino acid positions 134, 167, 198, or 200. However, two A. ceylanicum isolates from the Philippines harbored the novel V236I mutation resulting from a GTC → ATC nucleotide substitution (B). The mutation was identified through NGS-based sequence analysis of the aa198/200 amplicon region.
V236I mutation negatively affects BZ–β-tubulin interactions
To investigate the potential functional consequences of the novel V236I mutation, in silico structural biology approaches were employed to evaluate its effects on BZ–β-tubulin interactions. Molecular docking analyses demonstrated only minor alterations in docking poses and binding affinities between wild-type and mutant proteins (Figure 2A). Notably, the substitution did not produce major changes in the aa residues interacting directly with the ligand. Both the wild-type and mutant residues interacted with BZ ligands predominantly through weak van der Waals interactions.
Molecular dynamics simulations further demonstrated that β-tubulin harboring the V236I mutation exhibited dynamic behavior comparable to that of the wild-type protein and previously characterized Q134H and E198K mutants during interactions with ABZ and FBZ. The post-equilibration RMSD values of the V236I complexes (ABZ = 2.35 ± 0.07 Å; FBZ = 2.41 ± 0.09 Å) were comparable to those of the wild-type protein (ABZ = 2.42 ± 0.06 Å; FBZ = 2.21 ± 0.09 Å), the Q134H mutant (ABZ = 2.51 ± 0.08 Å; FBZ = 2.26 ± 0.08 Å), and the E198K mutant (ABZ = 2.26 ± 0.09 Å; FBZ = 2.52 ± 0.07 Å) (Figure 2B).
Similarly, RMSF analyses revealed highly comparable residue-level flexibility profiles among the wild-type and mutant β-tubulin structures, indicating that the V236I substitution exerts minimal influence on overall protein structural integrity (Figure 2C). Post-equilibration SASA values for the V236I mutant (ABZ = 1.86 ± 0.04 Ų; FBZ = 1.82 ± 0.03 Ų) were also comparable to those observed for the wild-type protein (ABZ = 1.87 ± 0.02 Ų; FBZ = 1.81 ± 0.03 Ų), the Q134H mutant (ABZ = 1.84 ± 0.02 Ų; FBZ = 1.82 ± 0.04 Ų), and the E198K mutant (ABZ = 1.78 ± 0.02 Ų; FBZ = 1.81 ± 0.02 Ų). These findings indicate that the V236I mutation does not substantially alter the dynamic behavior of β-tubulin during ligand binding and is unlikely to compromise the protein's structural stability or biological function.
Effect of V236I on binding free-energy and hydrogen-bond formation
Although the V236I mutation produced minimal effects on overall protein dynamics, molecular dynamics simulations predicted a weakening of the β-tubulin–BZ interaction. The energetic favorability of ligand-binding was assessed using MM-GBSA and MM-PBSA binding free-energy calculations based on equilibrated simulation trajectories (Supplementary Tables 1 and 2).
Overall, the V236I mutation reduced binding free energies compared with wild-type β-tubulin complexes (Figure 3A). This reduction was particularly pronounced when ABZ was used as the ligand. Interestingly, the observed binding-energy values were comparable to those previously reported for the Q134H and E198K mutants, both of which have been experimentally associated with BZ resistance. These findings suggest that the V236I mutation may similarly contribute to reduced susceptibility to BZ compounds, particularly ABZ.
To further investigate this observation, hydrogen-bond formation between β-tubulin and BZ ligands was examined. In general, the V236I mutant exhibited fewer hydrogen-bond interactions than the wild-type protein (Figure 3B). Per-residue energy decomposition analyses further demonstrated the contributions of individual aas to ligand binding (Figure 3C). All canonical resistance-associated positions (134, 167, 198, and 200) participated in ligand interactions.
When FBZ was used as the ligand, the overall interaction patterns remained largely conserved. Nevertheless, the substitution of valine by isoleucine at position 236 resulted in a reduction in residue-specific binding energy (wild-type: −2.66 kcal/mol; V236I: −1.25 kcal/mol). Likewise, when ABZ was used as the ligand, the overall pattern of interacting residues remained similar, although their individual energetic contributions were markedly diminished relative to the wild-type protein.
Collectively, these findings indicate that the V236I mutation is associated with reduced energetic favorability of β-tubulin–BZ interactions. The observed decrease in binding free-energy appears to be driven primarily by a reduction in hydrogen-bond formation between the mutant protein and the ligand, potentially compromising drug-binding efficiency.
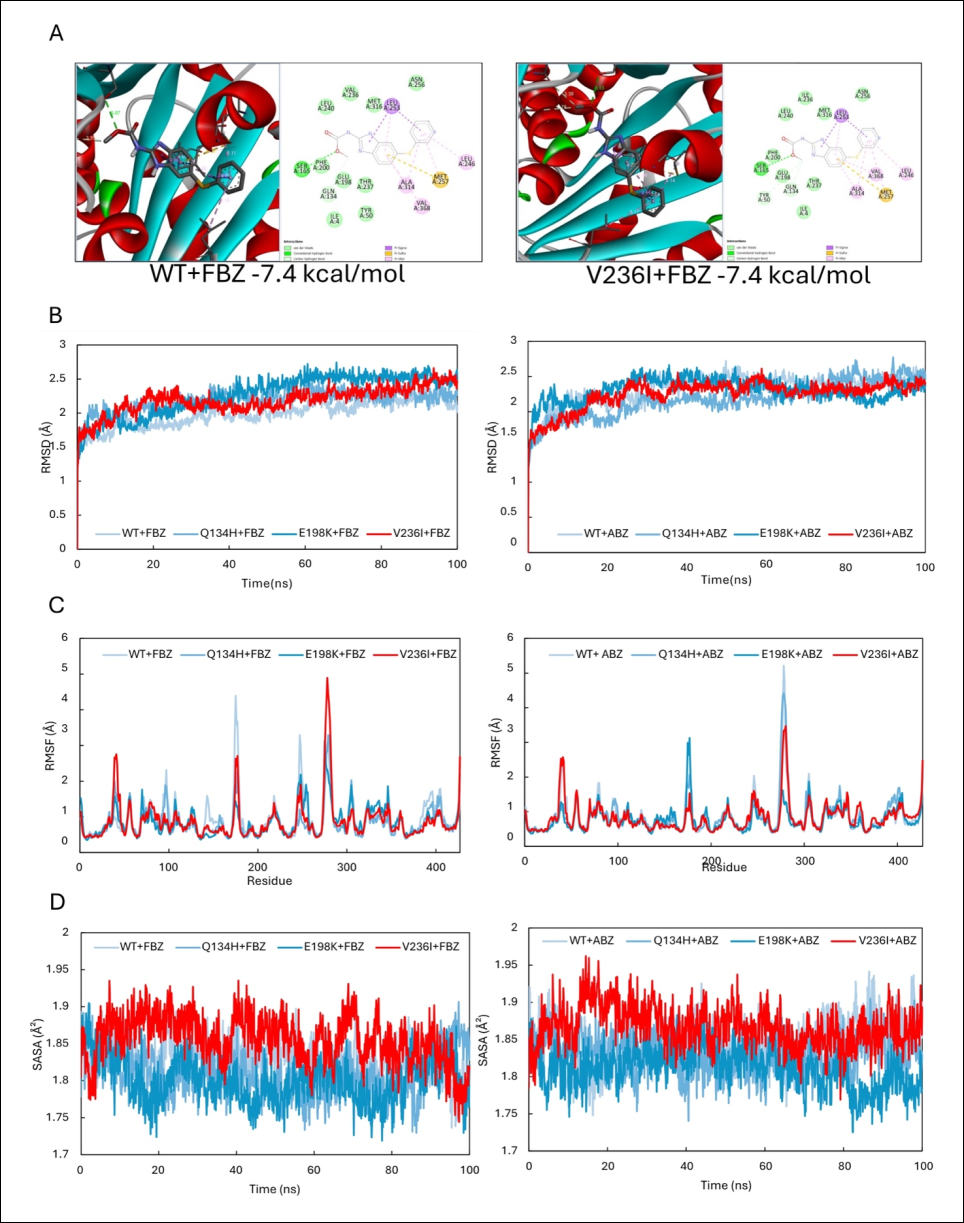
Figure 2. Molecular docking and molecular dynamics analyses of A. ceylanicum β-tubulin interactions with fenbendazole (FBZ) and albendazole (ABZ). Results corresponding to the V236I mutation are highlighted in red. (A) Docking poses and binding affinities obtained from molecular docking analyses. (B) RMSD profiles demonstrating that the dynamic binding behavior of β-tubulin–BZ complexes remained comparable among the wild-type protein and mutant variants regardless of the ligand used. (C) RMSF analyses showing that the V236I mutation did not substantially affect residue-level protein flexibility. (D) SASA analyses indicate that the mutation did not significantly alter the solvent-accessible surface area of the protein. Collectively, these findings suggest that the V236I mutation has minimal effects on the overall structural conformation and dynamic stability of hookworm β-tubulin.
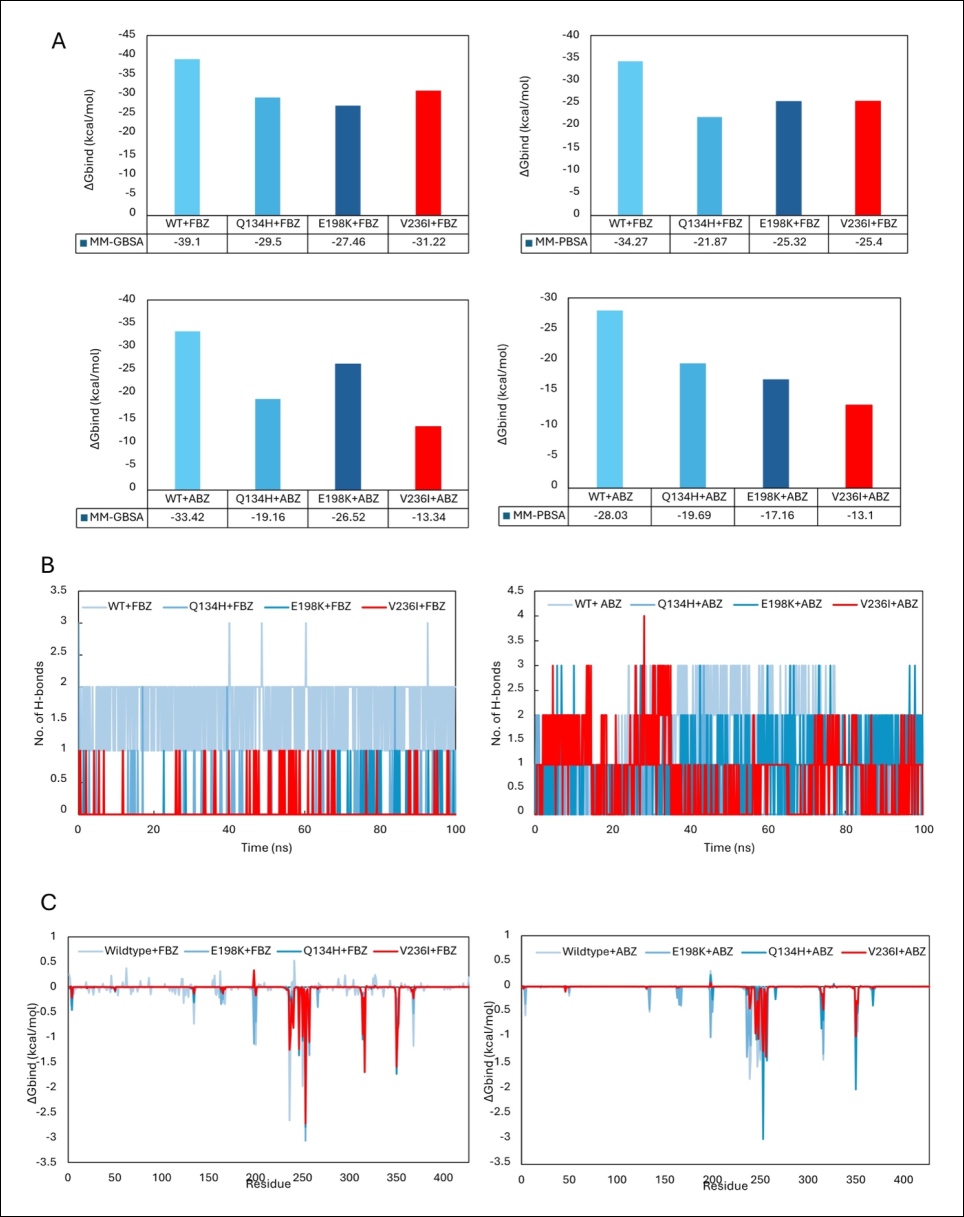
Figure 3. Interaction analyses between hookworm β-tubulin and BZ drugs. Results corresponding to the V236I mutation are highlighted in red. (A) MM-GBSA and MM-PBSA analyses demonstrated reduced binding free energies in the V236I mutant, indicating decreased energetic favorability of β-tubulin–BZ interactions. (B) The reduction in binding favorability was associated with a lower number of hydrogen-bond interactions between the mutant protein and BZ ligands. (C) Per-residue binding free-energy decomposition analyses illustrating the contributions of individual amino acid residues to ligand-binding and overall interaction energy.
V236I mutation has minimal effects on tubulin dimerization
The effects of the V236I mutation on tubulin dimerization were also investigated using in silico structural and molecular dynamics analyses. Results were compared with previously published data for wild-type A. ceylanicum tubulin dimers and dimers carrying the Q134H and E198K mutations [36].
Molecular dynamics simulations demonstrated that incorporation of the V236I mutation into the β-subunit did not substantially alter dimer behavior. The post-equilibration RMSD value of the V236I dimer (3.94 ± 0.19 Å) was comparable to those of the wild-type (3.75 ± 0.21 Å) and E198K (3.81 ± 0.17 Å) dimers and lower than that observed for the Q134H mutant (Figure 4A). Similar flexibility profiles were observed among all dimers, indicating preservation of the overall structural dynamics despite the presence of mutations (Figure 4B).
Likewise, SASA and radius of gyration analyses demonstrated comparable structural behavior among all evaluated dimers (Figures 4C and D). These findings indicate that the V236I mutation exerts minimal influence on tubulin dimer dynamics and maintains stable interactions between the α- and β-subunits.
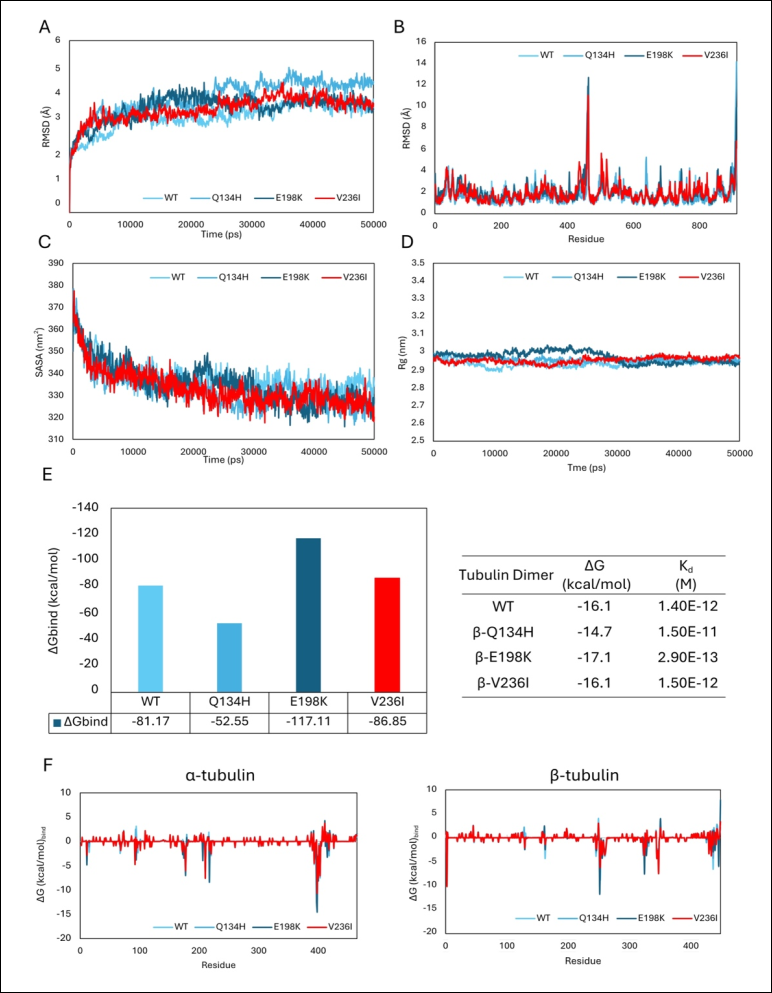
Figure 4. In silico analyses of hookworm tubulin dimerization. Results corresponding to the V236I mutation are highlighted in red. (A) RMSD profiles demonstrating stable dynamic behavior of both wild-type and mutant tubulin dimers throughout the simulation period. (B) RMSF analyses show similar flexibility patterns across all evaluated dimers, regardless of mutation status. (C) SASA analyses indicate comparable solvent-accessible surface areas between wild-type and mutant dimers. (D) Radius of gyration analyses demonstrated that the dimers remained structurally compact throughout the simulations. (E) MM-GBSA binding-energy estimations, binding affinities, and dissociation constants showed that the V236I mutant exhibited dimerization properties comparable to those of the wild-type protein. (F) Per-residue binding free-energy decomposition analyses demonstrate similar energetic contributions among amino acid residues in the mutant and wild-type dimers. Collectively, these findings indicate that the V236I mutation exerts minimal effects on tubulin dimerization and overall β-tubulin structural function.
Effect of V236I on dimer binding energetics
The energetic favorability of dimer formation was further assessed through MM-GBSA calculations. The V236I-containing dimer exhibited a slightly more favorable binding energy than the wild-type dimer (wild-type = −81.17 kcal/mol; V236I = −86.65 kcal/mol). Similarly, estimated binding affinity and dissociation constant values were highly comparable between mutant and wild-type dimers (Figure 4E).
Per-residue energy decomposition analyses also showed similar energetic contributions across aa residues, regardless of the presence of the V236I mutation (Figure 4F). Unlike the previously characterized E198K and Q134H mutations, which respectively increased and decreased dimer interaction favorability, the V236I mutation produced only negligible changes in dimer energetics.
Overall, these findings indicate that the novel V236I β-tubulin variant does not impair tubulin dimerization. The mutation does not reduce the energetic attraction between α- and β-subunits and is therefore unlikely to significantly alter the structural or functional properties of the tubulin dimer.
DISCUSSION
Occurrence of the novel V236I mutation in Southeast Asian hookworms
The present study investigated hookworm samples from the Philippines and Indonesia for evidence of BZ resistance by analyzing the β-tubulin isotype 1 gene. A novel aa substitution, V236I (valine → isoleucine), resulting from a SNP, was identified in two A. ceylanicum isolates from the Philippines. No canonical BZ resistance-associated mutations were detected in the analyzed samples. Furthermore, computational analyses demonstrated that the bulkier isoleucine side chain at position 236 may introduce steric hindrance within the ligand-binding region, thereby reducing drug-binding efficiency by decreasing hydrogen-bond formation and altering ligand positioning. Despite these effects on ligand-binding, the similar biochemical properties of valine and isoleucine appeared to minimize any substantial effects on tubulin dimerization.
Very few studies have investigated the occurrence of BZ resistance-associated mutations among STHs in Southeast Asia. Recently, Jones et al. [41] reported the global absence of BZ resistance-associated mutations in A. lumbricoides, including samples collected from the Philippines. In addition, a preprint study reported the emergence of BZ resistance-associated mutations in samples from Lao PDR using pyrosequencing techniques [42]. Mutations at codons 198 and 200 were detected in A. lumbricoides, T. trichiura, and N. americanus, whereas mutations at codon 167 were observed only in T. trichiura and N. americanus [42]. Importantly, the frequency of these mutations remained below 10% in the analyzed populations [42]. Among canine hookworms, BZ resistance-associated mutations have been reported to be widespread in A. caninum populations from Australia and New Zealand [19]. The F167Y and Q134H mutations predominated in those populations and were also detected in U. stenocephala for the first time. Moreover, allele frequencies exceeding 75% for F167Y were associated with marked reductions in BZ efficacy [19].
Our previous systematic review identified seven SNP-induced aa substitutions in the β-tubulin gene of hookworms associated with BZ resistance: Q134H, F167Y, E198A, E198K, E198V, F200Y, and F200L [22]. The emergence of the V236I mutation in the Philippines may be linked to the country's long-standing STH preventive chemotherapy program [43]. The Integrated Helminth Control Program of the Philippine Department of Health was initiated in 2006 and remains active today [44]. The program relies heavily on mass drug administration using BZ compounds among susceptible populations, particularly schoolchildren, alongside health education and water, sanitation, and hygiene interventions [29, 43].
The identification of the V236I mutation in a zoonotic hookworm species represents the first report of genetic variation at this position in the β-tubulin gene of hookworms in Southeast Asia. To the best of our knowledge, this is also the first report of a mutation at codon 236 in the β-tubulin gene of any helminth species. Consequently, its potential role as a candidate BZ resistance-associated variant warrants further epidemiological and functional investigation. The occurrence of this mutation in A. ceylanicum additionally highlights the zoonotic dimension of BZ resistance and reinforces the importance of a One Health approach for resistance surveillance and control [3,45]. Nevertheless, the relatively small sample size analyzed in this study should be considered when interpreting the findings.
Potential role of V236I in BZ resistance
Our in silico analyses indicate that the V236I mutation may play an important role in BZ binding due to its location within the ligand-binding region of β-tubulin. Similar to canonical resistance-associated mutations, this substitution results from a single-nucleotide change [7]. In the V236I variant, isoleucine replaces valine at an aa position located within the colchicine-binding region of the β-tubulin protein [46, 47].
In our previous study, we observed that the wild-type valine residue at position 236 participated in weak van der Waals interactions with BZ ligands when A. ceylanicum β-tubulin was used as the structural model [32]. Similarly, Jones et al. [46] reported that valine at the same position formed hydrogen-bond interactions with the amino group of the imidazole ring of the BZ core in the presence of the F200Y mutation in A. duodenale β-tubulin. Furthermore, analyses of wild-type and mutant ascarid β-tubulins demonstrated that this residue participates in hydrogen-bond, π-alkyl, and van der Waals interactions with BZ ligands [46].
Collectively, these observations suggest that aa position 236 contributes to the stabilization of BZ–β-tubulin interactions. The interactions mediated by this residue may complement the critical interactions formed by aas located at canonical resistance-associated positions, particularly glutamine at position 198 [47, 48]. Therefore, mutations occurring at this site may have functional consequences for drug-binding even though they are located outside the traditionally recognized resistance-associated codons.
Potential steric and energetic mechanisms underlying reduced BZ-binding
Our in silico analyses suggest that the V236I mutation may impair BZ-binding, potentially through both steric and energetic mechanisms. The substitution of valine with isoleucine resulted in reduced binding free-energy between β-tubulin and BZ ligands. In addition, a reduction in hydrogen-bond formation was observed, indicating a less favorable interaction between the mutant protein and the drug molecule.
These findings are consistent with our previous analyses of the Q134H and E198K mutations, both of which demonstrated reduced BZ-binding favorability and have been experimentally associated with BZ resistance [9, 18, 34]. In those variants, alterations in aa functional groups modified the biochemical characteristics of the ligand-binding region. For example, replacement of glutamic acid with lysine changes the local charge environment because glutamic acid carries a negatively charged carboxyl group, whereas lysine possesses a positively charged amino group [46, 47]. Furthermore, lysine at position 198 disrupts hydrogen-bond interactions with the imidazole ring of the BZ core, which are essential for efficient drug-binding [46, 47, 49].
Unlike E198K, the V236I mutation does not substantially alter the biochemical properties of the protein because both valine and isoleucine possess nonpolar aliphatic side chains. Structurally, however, isoleucine contains a bulkier sec-butyl side-chain compared with the isopropyl side-chain of valine. This increase in side-chain volume may create steric hindrance within the binding pocket [46]. Consequently, the additional steric bulk introduced by isoleucine may interfere with optimal ligand positioning and reduce binding efficiency.
Consistent with this hypothesis, we observed reductions in both binding-site volume and binding-site area when the V236I mutation was present (Supplementary Figure 5). Although these structural changes were relatively modest, they may be sufficient to displace the ligand from its optimal binding orientation, thereby reducing hydrogen-bond formation and the overall binding free energy. The ligand RMSD trajectories generated during molecular dynamics simulations support this interpretation, particularly for ABZ-bound complexes (Supplementary Figure 7).
Evidence from other biological systems further supports this mechanism. For example, substitution of alanine by valine at position 2004 of acetyl-CoA carboxylase in Alopecurus myosuroides reduced susceptibility to the herbicide Quizalofop [50]. The introduction of a bulkier side-chain disrupted optimal ligand interactions and reduced herbicidal efficacy. Our findings suggest that a comparable steric hindrance mechanism may occur when the V236I mutation is present in hookworm β-tubulin. Nevertheless, additional validation through in vitro and in vivo studies, including gene-editing approaches similar to those employed by Venkatesan et al. [18], is required to confirm the biological significance of this mutation.
Effect of V236I on tubulin dimerization and structural stability
To evaluate whether the V236I mutation influences microtubule assembly, we investigated its effects on tubulin dimerization using multiple in silico approaches. The results demonstrated that the mutation exerts minimal influence on dimer dynamics, binding efficiency, and overall structural stability. These findings contrast with our previous work examining the effects of other BZ resistance-associated mutations on tubulin dimerization using the same analytical framework [34].
The substitution of valine by isoleucine produced only minor alterations in the structural and functional characteristics of β-tubulin, which likely explains the limited effects observed on dimerization. In contrast, the E198K and Q134H mutations exhibited opposing effects on tubulin dimer stability, with E198K enhancing and Q134H reducing dimer binding efficiency [36]. These observations suggest that different resistance-associated mutations may contribute to resistance through distinct structural mechanisms.
Borchert et al. [48] reported that the presence of BZ compounds and resistance-associated mutations such as F167Y, E198A, and F200Y altered the lengths and rotational angles of β-sheets 4 and 6 within β-tubulin. The V236I mutation occurs within α-helix 7, a secondary structural element that has also been reported to undergo conformational changes in the presence of resistance-associated mutations [48]. However, the present findings suggest that any conformational alterations associated with V236I are likely minimal.
Furthermore, E198K has been reported to favorably influence the conformation of α-helix 8 and loop 8, regions containing major β-subunit binding residues that interact with loop 3 of the α-tubulin subunit [36]. Such favorable conformational effects were not observed for the V236I mutation, which may explain why the energetic and structural properties of the V236I-containing dimer closely resembled those of the wild-type dimer.
Taken together, the findings of the present study, our previous investigations, and published literature indicate that BZ resistance-associated mutations may influence tubulin dimer formation through structural alterations of the β-subunit. However, because of the high biochemical similarity between valine and isoleucine, the V236I mutation appears to exert only negligible effects on tubulin dimerization and overall dimer stability.
Study limitations and future research directions
Several limitations should be considered when interpreting the findings of this study. First, the number of hookworm samples analyzed was relatively small, which may limit the generalizability of the observed mutation frequencies. Second, the functional implications of the V236I mutation were inferred primarily from computational analyses rather than direct biological experiments. Although molecular docking and molecular dynamics simulations provide valuable mechanistic insights, they cannot fully replicate the complexity of biological systems.
Future studies should therefore investigate the phenotypic consequences of the V236I mutation through in vitro drug-susceptibility assays, gene-editing approaches, and in vivo validation experiments. Larger-scale molecular surveillance studies across Southeast Asia are also needed to determine the prevalence, geographic distribution, and evolutionary dynamics of this novel variant. Such investigations will be essential for determining whether V236I represents an emerging BZ resistance-associated marker in hookworm populations.
CONCLUSION
This study represents the first investigation of BZ resistance-associated mutations in hookworms from the Philippines and Indonesia using molecular characterization and comprehensive in silico analyses of the β-tubulin isotype 1 gene. A novel V236I aa substitution resulting from a GTC → ATC nucleotide change was identified in two A. ceylanicum isolates from the Philippines. No canonical BZ resistance-associated mutations were detected at codons 134, 167, 198, or 200. Computational analyses demonstrated that the V236I mutation was associated with reduced BZ-binding favorability, particularly for ABZ, as evidenced by decreased binding free energies, reduced hydrogen-bond formation, and alterations in ligand-binding behavior. These findings suggest that the mutation may weaken BZ–β-tubulin interactions through steric and energetic mechanisms arising from the bulkier isoleucine side-chain.
Despite its apparent effects on ligand binding, the V236I mutation had minimal effects on tubulin dimerization, structural stability, and conformational dynamics. Molecular dynamics simulations demonstrated that the mutant protein retained RMSD, RMSF, SASA, binding affinity, and dimer interaction profiles comparable to those of the wild-type protein. These observations indicate that the mutation may reduce drug-binding efficiency while preserving the essential structural and functional properties of β-tubulin required for parasite survival.
The identification of a previously unreported mutation in a zoonotic hookworm species has important practical implications for helminth control programs. Given the extensive use of BZ compounds in preventive chemotherapy programs throughout Southeast Asia, the emergence of novel variants that may reduce drug susceptibility highlights the need for routine molecular surveillance and early detection of resistance-associated markers. The occurrence of this mutation in A. ceylanicum further emphasizes the importance of adopting a One Health perspective when monitoring the emergence and spread of anthelmintic resistance.
A major strength of this study is the integration of molecular epidemiology with multiple complementary in silico approaches, including molecular docking, molecular dynamics simulations, binding free-energy calculations, and tubulin dimerization analyses. Together, these methods provided mechanistic insights into the potential biological significance of the newly identified mutation.
In conclusion, the novel V236I substitution identified in A. ceylanicum represents a promising candidate BZ resistance-associated variant that may reduce drug-binding efficiency while preserving tubulin structural integrity. These findings provide new insights into the molecular basis of potential BZ resistance in hookworms and underscore the importance of continued surveillance, functional validation, and proactive resistance-monitoring strategies to safeguard the long-term effectiveness of BZ-based control programs.
DATA AVAILABILITY
Raw sequences are available at NCBI BioProject Accession No. PRJNA1451827. In silico modeling data and results are available on Zenodo (https://doi.org/10.5281/zenodo.19499223).
AUTHORS’ CONTRIBUTIONS
JCB: Conceptualization, Formal analysis, investigation, writing – original draft, and writing – review and editing. IKBT, JMA, and MMMK: Investigation and writing – review and editing. MABM and FLO: Formal analysis and writing – review and editing. SS and PS: Conceptualization, supervision, and writing – review and editing. All authors have read and approved the final manuscript.
COMPETING INTERESTS
The authors declare that they have no competing interests.
PUBLISHER’S NOTE
Veterinary World (Publisher of International Journal of One Health) remains neutral with regard to jurisdictional claims in the published institutional affiliations.
ACKNOWLEDGMENTS
This study was supported by the Invitation Research Grant of the Faculty of Medicine, Khon Kaen University, Thailand (Grant Number: IN68071).
REFERENCES
- Ng'etich AI, Amoah ID, Bux F, Kumari S. Anthelmintic resistance in soil-transmitted helminths: One Health considerations. Parasitol Res 2023;123:62. [Google Scholar]
- Keiser J. Present drugs and future perspectives in treating soil-transmitted helminthiasis. Front Trop Dis 2023. [Google Scholar]
- Tenorio JCB, Tabios IKB, Inpankaew T, Ybañez AP, Tiwananthagorn S, Tangkawattana S, Suttiprapa S. Ancylostoma ceylanicum and other zoonotic canine hookworms: Neglected public and animal health risks in the Asia-Pacific region. Anim Dis 2024;4:11. [Google Scholar]
- Pattanayak P, Panigrahi D, Kumari S, Yadav HN, Ashby CR, Kerber S, Shahwan MJSA, Tiwari AK, Mishra GP. Benzimidazoles in helminthiasis chemotherapy: Developments and challenges. J Mol Struct 2024;1295:136716. [Google Scholar]
- Monga J, Ghosh NS, Rani I, Singh R, Deswal G, Dhingra AK, Grewal AS. Unlocking the pharmacological potential of benzimidazole derivatives: A pathway to drug development. Curr Top Med Chem 2024;24:437-485. [Google Scholar]
- Whittaker JH, Carlson SA, Jones DE, Brewer MT. Molecular mechanisms for anthelmintic resistance in strongyle nematode parasites of veterinary importance. J Vet Pharmacol Ther 2017;40(2):105-115. [Google Scholar]
- Furtado LFV, Paiva Bello ACP, Rabelo ÉML. Benzimidazole resistance in helminths: From problem to diagnosis. Acta Trop 2016;162:95-102. [Google Scholar]
- Von Samson-Himmelstjerna G, Blackhall WJ, McCarthy JS, Skuce PJ. Single-nucleotide polymorphism (SNP) markers for benzimidazole resistance in veterinary nematodes. Parasitology 2007;134(8):1077-1086. [Google Scholar]
- Dilks CM, Hahnel SR, Sheng Q, Long L, McGrath PT, Andersen EC. Quantitative benzimidazole resistance and fitness effects of parasitic nematode beta-tubulin alleles. Int J Parasitol Drugs Drug Resist 2020;14:28-36. [Google Scholar]
- Dilks CM, Koury EJ, Buchanan CM, Andersen EC. Newly identified parasitic nematode beta-tubulin alleles confer resistance to benzimidazoles. Int J Parasitol Drugs Drug Resist 2021;17:168-175. [Google Scholar]
- Jimenez Castro PD, Howell SB, Schaefer JJ, Avramenko RW, Gilleard JS, Kaplan RM. Multiple drug resistance in the canine hookworm Ancylostoma caninum: An emerging threat? Parasit Vectors 2019;12:576. [Google Scholar]
- Diawara A, Halpenny CM, Churcher TS, Mwandawiro C, Kihara J, Kaplan RM, Streit TG, Idaghdour Y, Scott ME, Basáñez MG, Prichard RK. Association between response to albendazole treatment and β-tubulin genotype frequencies in soil-transmitted helminths. PLoS Negl Trop Dis 2013;7(5):e2247. [Google Scholar]
- George S, Suwondo P, Akorli J, Otchere J, Harrison LM, Bilguvar K, Knight JR, Humphries D, Wilson MD, Caccone A, Cappello M. Application of multiplex amplicon deep-sequencing (MAD-seq) to screen for putative drug resistance markers in the Necator americanus isotype-1 β-tubulin gene. Sci Rep 2022;12:11459. [Google Scholar]
- Jimenez Castro PD, Venkatesan A, Redman E, Chen R, Malatesta A, Huff H, Zuluaga Salazar DA, Avramenko R, Gilleard JS, Kaplan RM. Multiple drug resistance in hookworms infecting greyhound dogs in the USA. Int J Parasitol Drugs Drug Resist 2021;17:107-117. [Google Scholar]
- Jimenez Castro PD, Kaplan RM. Persistent or suspected-resistant hookworm infections. Clin Brief 2020;18:59-68. [Google Scholar]
- Evason MD, Weese JS, Polansky B, Leutenegger CM. Emergence of canine hookworm treatment resistance: Novel detection of Ancylostoma caninum anthelmintic resistance markers by fecal PCR in 11 dogs from Canada. Am J Vet Res 2023. [Google Scholar]
- Leutenegger CM, Evason MD, Willcox JL, Rochani H, Richmond HL, Meeks C, Lozoya CE, Tereski J, Loo S, Mitchell K, Andrews J, Savard C. Benzimidazole F167Y polymorphism in the canine hookworm, Ancylostoma caninum: Widespread geographic, seasonal, age, and breed distribution in United States and Canada dogs. Int J Parasitol Drugs Drug Resist 2024;24:100520. [Google Scholar]
- Venkatesan A, Castro PDJ, Morosetti A, Horvath H, Chen R, Redman E, Dunn K, Collins JB, Fraser JS, Andersen EC, Kaplan RM, Gilleard JS. Molecular evidence of widespread benzimidazole drug resistance in Ancylostoma caninum from domestic dogs throughout the USA and discovery of a novel β-tubulin benzimidazole resistance mutation. PLoS Pathog 2023;19(8):e1011146. [Google Scholar]
- Abdullah S, Stocker T, Kang H, Scott I, Hayward D, Jaensch S, Ward MP, Jones MK, Kotze AC, Šlapeta J. Widespread occurrence of benzimidazole resistance single-nucleotide polymorphisms in the canine hookworm, Ancylostoma caninum, in Australia. Int J Parasitol 2024. [Google Scholar]
- Zuccherato LW, Furtado LFV, Medeiros CS, Pinheiro CS, Rabelo ÉM. PCR-RFLP screening of polymorphisms associated with benzimidazole resistance in Necator americanus and Ascaris lumbricoides from different geographical regions in Brazil. PLoS Negl Trop Dis 2018;12(10):e0006766. [Google Scholar]
- Grau-Pujol B, Gandasegui J, Escola V, Marti-Soler H, Cambra-Pellejà M, Demontis M, Brienen EAT, Jamine JC, Muchisse O, Cossa A, Sacoor C, Cano J, Lieshout LV, Martinez-Valladares M, Muñoz J. Single-nucleotide polymorphisms in the beta-tubulin gene and its relationship with treatment response to albendazole in human soil-transmitted helminths in southern Mozambique. Am J Trop Med Hyg 2022;107(3):649-657. [Google Scholar]
- Tenorio JCB, Heikal MF, Kafle A, Saichua P, Suttiprapa S. Benzimidazole resistance-associated mutations in the β-tubulin gene of hookworms: A systematic review. Parasitol Res 2024;123:405. [Google Scholar]
- Tinkler SH. Preventive chemotherapy and anthelmintic resistance of soil-transmitted helminths – Can we learn nothing from veterinary medicine? One Health 2020;9:100106. [Google Scholar]
- Emerson PM, Evans D, Freeman MC, Hanson C, Kalua K, Keiser J, Krolewiecki A, Leonard L, Levecke B, Matendechero S, Means AR, Montresor A, Mupfasoni D, Pullan RL, Rotondo LA, Stephens M, Sullivan KM, Walson JL, Williams T, Utzinger J. Need for a paradigm shift in soil-transmitted helminthiasis control: Targeting the right people, in the right place, and with the right drug(s). PLoS Negl Trop Dis 2024;18:e0012521. [Google Scholar]
- Colella V, Khieu V, Worsley A, Senevirathna D, Muth S, Huy R, Odermatt P, Traub RJ. Risk profiling and efficacy of albendazole against the hookworms Necator americanus and Ancylostoma ceylanicum in Cambodia to support control programs in Southeast Asia and the Western Pacific. Lancet Reg Health West Pac 2021;16:100258. [Google Scholar]
- Hong S, Jung BK, Song H, Cho J, Sohn WM, Hong SJ, Ryu JY, Kim HS, Jeong MH, Htoon TT, Tin HH, Chai JY. Failure of repeated MDA with albendazole for trichuriasis control in schoolchildren of the Yangon Region, Myanmar. Korean J Parasitol 2021;59(6):607-613. [Google Scholar]
- Becker SL, Liwanag HJ, Snyder JS, Akogun O, Jr VB, Freeman MC, Gyorkos TW, Imtiaz R, Keiser J, Krolewiecki A, Levecke B, Mwandawiro C, Pullan RL, Addiss DG, Utzinger J. Toward the 2020;12:e0006606. [Google Scholar]
- Jex AR, Lim YAL, Bethony JM, Hotez PJ, Young ND, Gasser RB. Soil-transmitted helminths of humans in Southeast Asia—Towards integrated control. Adv Parasitol 2011;74:231-265. [Google Scholar]
- Mationg MLS, Tallo VL, Williams GM, Gordon CA, Clements ACA, McManus DP, Gray DJ. The control of soil-transmitted helminthiases in the Philippines: The story continues. Infect Dis Poverty 2021;10:85. [Google Scholar]
- Traub RJ, Inpankaew T, Sutthikornchai C, Sukthana Y, Thompson RCA. PCR-based coprodiagnostic tools reveal dogs as reservoirs of zoonotic ancylostomiasis caused by Ancylostoma ceylanicum in temple communities in Bangkok. Vet Parasitol 2008;155(1–2):67-73. [Google Scholar]
- Medeiros CDS, Furtado LFV, Miranda GS, Silva VJ, Santos TR, Rabelo ÉML. Moving beyond the state of the art of understanding resistance mechanisms in hookworms: Confirming old and suggesting new associated SNPs. Acta Trop 2022;233:106533. [Google Scholar]
- Tenorio JCB, Heikal MF, Kafle A, Macalalad MAB, Orosco FL, Saichua P, Suttiprapa S. Unraveling the mechanisms of benzimidazole resistance in hookworms: A molecular docking and dynamics study. J Genet Eng Biotechnol 2025;23:100472. [Google Scholar]
- Trott O, Olson AJ. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem 2010;31(2):455-461. [Google Scholar]
- Tenorio JCB, Heikal MF, Kafle A, Saichua P, Suttiprapa S. Benzimidazole resistance-associated mutations improve the in silico dimerization of hookworm tubulin: An additional resistance mechanism. Vet World 2024;17(11):2736-2746. [Google Scholar]
- Abramson J, Adler J, Dunger J, Evans R, Green T, Pritzel A, Ronneberger O, Willmore L, Ballard AJ, Bambrick J, Bodenstein SW, Evans DA, Hung CC, O'Neill M, Reiman D, Tunyasuvunakool K, Wu Z, Žemgulytė A, Arvaniti E, Beattie C, Bertolli O, Bridgland A, Cherepanov A, Congreve M, Cowen-Rivers AI, Cowie A, Figurnov M, Fuchs FB, Gladman H, Jain R, Khan YA, Low CMR, Perlin K, Potapenko A, Savy P, Singh S, Stecula A, Thillaisundaram A, Tong C, Yakneen S, Zhong ED, Zielinski M, Žídek A, Bapst V, Kohli P, Jaderberg M, Hassabis D, Jumper JM. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024;630(8016):493-500. [Google Scholar]
- Nogales E. Structural insights into microtubule function. Annu Rev Biochem 2000;69:277-302. [Google Scholar]
- Heo L, Lee H, Seok C. GalaxyRefineComplex: Refinement of protein–protein complex model structures driven by interface repacking. Sci Rep 2016;6:32153. [Google Scholar]
- eng G, Wang E, Wang Z, Liu H, Zhu F, Li D, Hou T. HawkDock: A web server to predict and analyze the protein–protein complex based on computational docking and MM/GBSA. Nucleic Acids Res 2019;47(W1):W322-W330. [Google Scholar]
- Jones BP, van Vliet AHM, LaCourse EJ, Betson M. In silico docking of nematode β-tubulins with benzimidazoles points to gene expression and orthologue variation as factors in anthelmintic resistance. Front Trop Dis 2022;3:898814. [Google Scholar]
- Robinson MW, McFerran N, Trudgett A, Hoey L, Fairweather I. A possible model of benzimidazole-binding to beta-tubulin disclosed by invoking an inter-domain movement. J Mol Graph Model 2004;23(3):275-284. [Google Scholar]
- Jones BP, Kozel K, Alonte AJI, Llanes KKR, Juhász A, Chaudhry U, Roose S, Geldhof P, Belizario VY, Nejsum P, Stothard JR, LaCourse EJ, van Vliet AHM, Paller VGV, Betson M. Worldwide absence of canonical benzimidazole resistance-associated mutations within β-tubulin genes from Ascaris. Parasit Vectors 2024;17:225. [Google Scholar]
- Tenorio JCB, Belizario VY Jr, Furtado LFV, Suttiprapa S. Can benzimidazole resistance undermine the Philippines' success in controlling and eliminating soil-transmitted helminth infections? A mini-review. Infect Microbes Dis 2024. [Google Scholar]
- Philippine Department of Health. Administrative Order 2006. [Google Scholar]
- Can resistant Ancylostoma ceylanicum arise and spread between humans and animals?. Microbes Infect Dis 2025;6(4):6864-6867. [Google Scholar]
- Jones BP, van Vliet AHM, LaCourse EJ, Betson M. Identification of key interactions of benzimidazole resistance-associated amino acid mutations in Ascaris β-tubulins by molecular docking simulations. Sci Rep 2022;12:13725. [Google Scholar]
- Aguayo-Ortiz R, Méndez-Lucio O, Romo-Mancillas A, Castillo R, Yépez-Mulia L, Medina-Franco JL, Hernández-Campos A. Molecular basis for benzimidazole resistance from a novel β-tubulin binding site model. J Mol Graph Model 2013;45:26-37. [Google Scholar]
- Borchert M, Hellinga JR, Reber S, Krücken J, Von Samson-Himmelstjerna G. Benzimidazole inhibits Haemonchus contortus microtubule dynamics by intradimer structural changes observed by in silico modeling. J Biomol Struct Dyn 2024. [Google Scholar]
- Mann A. Conformational restriction and/or steric hindrance in medicinal chemistry. In: Wermuth CG, editor. The Practice of Medicinal Chemistry. 3rd ed. New York: Academic Press; 2008. [Google Scholar]
- Bough R, Dayan FE. Biochemical and structural characterization of quizalofop-resistant wheat acetyl-CoA carboxylase. Sci Rep 2022;12:679. [Google Scholar]